instituto de biologia molecular e celular | institute for molecular and cell biology
![]()
Projecto nº:016589
Referência do Projecto:PTDC/BEX-BCM/1758/2014 (POCI-01-0125-FEDER-016589)
Título:Spatiotemporal Control of Mitotic Spindle Assembly.
Montante envolvidos:
Investimento total: 199.824,00 €
IBMC-Instituto de Biologia Molecular e Celular -183.768,00€
Apoio FEDER: 156.202,80€
Apoio OE: 27.565,20€
INEB-Instituto Nacional de Engenharia Biomédica -16.056,00€
Apoio FEDER: 13.647,60€
Apoio OE: 2.408,40€
Localização do projecto: Porto, Portugal
Sintese do projecto:
Mitosis is the process by which eukaryotic cells transmit genetic information to the following generation. Therefore, itis not surprising that errors during mitosis can lead to aneuploidy and genetic instability and, consequently, have been linked to developmental abnormalities and cancer progression. Clearly, uncovering the basic mechanisms controlling mitotic fidelity will have strong implications for our understanding of carcinogenesis.
For chromosome segregation to occur, cells need to assemble a microtubule (MT)-based structure known as mitotic spindle. Although there are multiple pathways contributing to spindle assembly [1, 2], in human cells this process relies mainly on centrosomes (CTs), the main MT-organizing centre. In preparation for mitosis, duplicated CTs need to separate along the nuclear envelope. Multiple players have been involved in the process, including molecular motors such as Eg5 and Dynein, as well as actin, myosin II and MTs [3-7]. However, how these different components are functionally coordinated remains unclear. More specifically, we do not know how the force generated by these different players can be directed in such a way that CTs migrate to opposite sides of the nucleus. This knowledge is of crucial importance when we consider that CTs can affect mitotic fidelity. In fact, the role of CTs on mitotic fidelity and aneuploidy has been identified as early as the 19th century with the works of Boveri and van Beneden. In addition, it has also been shown that mitotic progression relies on the correct separation of CTs prior to Nuclear Envelope Breakdown (NEB) [8, 9]. Failure to do so results in the occurrence of erroneous attachments between MTs and a structure on the chromosome known as kinetochore (KT) [9]. Some of these errors include merotelic attachments, which involve simultaneous binding of MTs from opposite poles to the same KT, therefore satisfying the Spindle
Assembly Checkpoint (SAC) that monitors KT-MT attachment, but causing lagging chromosomes during anaphase.
Therefore, defining the combined contributions of all molecular players involved will be crucial to understanding the causal relationship between CT separation and mitotic fidelity.
Pioneering work from Hertwig in 1884, allowed the establishment of a simple set of rules relating cell shape with the division axis. He proposed that 1) the nucleus tends to take up a position on the centre of its sphere of influence i.e. of the protoplasmic mass in which it lies? and 2) ?the axis of the mitotic figures typically lies in the longest axis of the protoplasmic mass, and division therefore tends to cut this at a right angle. These observations indicate that nuclear position inside the cell is not random and that cells retain spatial information relevant for spindle positioning, so that spindles align with the long cell axis (known as the ?Hertwig Rule?). Interestingly, recent data from one of our team members showed that this is a dynamic process and that spindle positioning responds to forces exerted from the extracellular environment [10]. Although metaphase spindles (and consequently CTs) are usually aligned with the long cell axis, little is known about how the imposition of spatial and geometrical constraints affects CT separation and early spindle assembly. Importantly, it is not known whether and how these constraints affect mitotic fidelity. Interestingly,
a recent report demonstrated that changes in cell geometry, such as those observed during mitotic cell rounding, are required for an efficient spindle assembly [11]. Whether these changes affect spindle assembly by interfering with timely and correct CT separation remains to be determined.
Here we propose a multi-disciplinary project to dissect, from a molecular perspective, how the interaction between the cell cortex, CTs and nucleus controls early spindle assembly and mitotic fidelity in space and time. To do this we will undertake several parallel approaches: a) thorough and quantitative spatiotemporal analysis of CT behaviour during mitosis using advanced micropatterning and cell confinement technology; b) use of state-of-the-art live cell microscopy and laser microsurgery combined with RNAi techniques to functionally dissect the role of the players involved in CT separation, spindle assembly and mitotic fidelity; and c) development of a biophysical model to assess the possible contributions of nuclear shape and CT position to spindle assembly efficiency.
Combining these experimental and computational approaches, we expect to determine which are the physical and molecular factors controlling CT behaviour and early spindle assembly. Moreover, we will obtain detailed information on how changing cellular geometry affects mitotic progression. Ultimately, we aim to determine how CT separation influences chromosome segregation and eventually, establish the basis of a new regulatory mechanism that ensures mitotic fidelity by actively monitoring cellular shape.
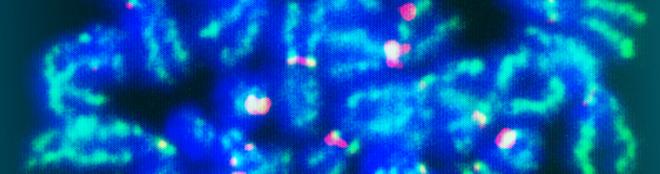
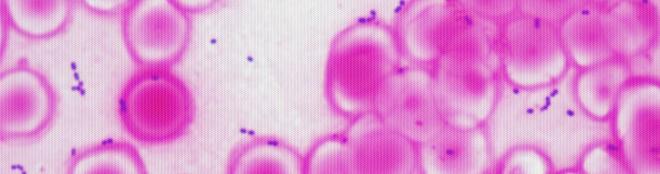
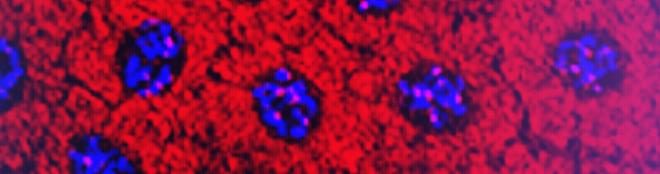
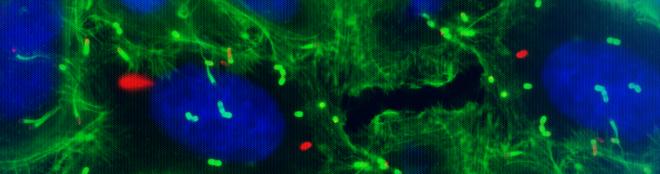
Galeria de fotos do projeto
Home | Site Map | Contacts | Credits | Privacy & Cookies | WHISTLEBLOWER CHANNEL | Intranet | Social Networks |

rua alfredo allen, 208, 4200-135 porto - portugal | tel +351 226 074 900 | email: info@i3s.up.pt | © copyright 2010 ibmc ![]()